Caspase 8 抗体
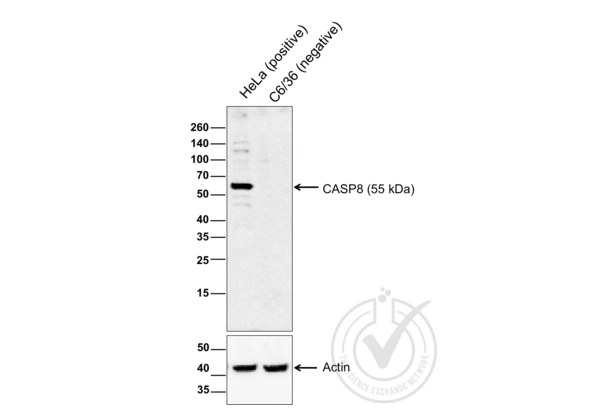 Caspase 8 抗体 (AA 411-482) (ABIN724205)
Caspase 8 抗体 (AA 411-482) (ABIN724205)
CASP8 适用: 人, 小鼠, 大鼠, 猴, Pig, 绵羊 WB, ELISA, IHC (p), FACS, IF (cc), IF (p), IHC (fro) 宿主: 兔 Polyclonal unconjugated
CASP8 适用: 人, 小鼠, 大鼠 WB, ELISA, IHC (p), FACS, IF (cc), IF (p), IHC (fro) 宿主: 兔 Polyclonal unconjugated
Caspase 8 抗体 by Grade
Find Caspase 8 抗体 with a specific Grade. The Grade listed below are among those available. Click on a link to go to the corresponding products.
Caspase 8 抗体 by 适用
Find Caspase 8 抗体 for a variety of species such as anti-Human Caspase 8, anti-Mouse Caspase 8, anti-Rat Caspase 8. The species listed below are among those available. Click on a link to go to the corresponding products.
Caspase 8 抗体 by 抗体来源
Find Caspase 8 抗体 with a specific 抗体来源. The 抗体来源 listed below are among those available. Click on a link to go to the corresponding products.
Caspase 8 抗体 by 克隆形成能力
Find available monoclonal or polyclonal Caspase 8 抗体. Click on a link to go to the corresponding products.
Popular Caspase 8 抗体
- (15)
- (5)
- (1)
- (17)
- (12)
- (4)
- (5)
- (10)
- (9)
- (9)
- (2)
- (5)
- (9)
- (6)
- (4)
- (4)
- (4)
- (4)
- (3)
- (3)
- (3)
Latest Publications for our Caspase 8 抗体
: "NLRP12 collaborates with NLRP3 and NLRC4 to promote pyroptosis inducing ganglion cell death of acute glaucoma." in: Molecular neurodegeneration, Vol. 15, Issue 1, pp. 26, (2020) (PubMed).: "The protective role of the MKP-5-JNK/P38 pathway in glucolipotoxicity-induced islet β-cell dysfunction and apoptosis." in: Experimental cell research, Vol. 382, Issue 1, pp. 111467, (2020) (PubMed).
: "Downregulation of METTL14 increases apoptosis and autophagy induced by cisplatin in pancreatic cancer cells." in: The international journal of biochemistry & cell biology, Vol. 122, pp. 105731, (2020) (PubMed).
: "Triggering apoptosis by oroxylin A through caspase-8 activation and p62/SQSTM1 proteolysis." in: Redox biology, Vol. 29, pp. 101392, (2020) (PubMed).
: "Emodin induced necroptosis in the glioma cell line U251 via the TNF-α/RIP1/RIP3 pathway." in: Investigational new drugs, Vol. 38, Issue 1, pp. 50-59, (2020) (PubMed).
: "Microglia-derived TNF-α mediates endothelial necroptosis aggravating blood brain-barrier disruption after ischemic stroke." in: Cell death & disease, Vol. 10, Issue 7, pp. 487, (2020) (PubMed).
: "GL-V9 exerts anti-T cell malignancies effects via promoting lysosome-dependent AKT1 degradation and activating AKT1/FOXO3A/BIM axis." in: Free radical biology & medicine, Vol. 145, pp. 237-249, (2020) (PubMed).
: "Xanthones from the Bark of Garcinia xanthochymus and the Mechanism of Induced Apoptosis in Human Hepatocellular Carcinoma HepG2 Cells via the Mitochondrial Pathway." in: International journal of molecular sciences, Vol. 20, Issue 19, (2020) (PubMed).
: "Histone methyltransferase SETDB1 promotes colorectal cancer proliferation through the STAT1-CCND1/CDK6 axis." in: Carcinogenesis, Vol. 41, Issue 5, pp. 678-688, (2020) (PubMed).
: "Giardia duodenalis induces extrinsic pathway of apoptosis in intestinal epithelial cells through activation of TNFR1 and K63 de-ubiquitination of RIP1 in vitro." in: Microbial pathogenesis, Vol. 149, pp. 104315, (2020) (PubMed).
Aliases for Caspase 8 抗体
caspase 8 L homeolog (casp8.L) 抗体death related ced-3/Nedd2-like protein (LOC661095) 抗体
caspase 8 (CASP8) 抗体
caspase-8 (LOC5563818) 抗体
caspase-8 (casp8) 抗体
caspase-8-like (LOC100222284) 抗体
caspase 8 (Casp8) 抗体
caspase 8, apoptosis-related cysteine peptidase (casp8) 抗体
ALPS2B 抗体
CAP4 抗体
CASP-8 抗体
Casp-8 抗体
casp8 抗体
CASP8 抗体
casp8-A 抗体
caspase-8 抗体
FLICE 抗体
MACH 抗体
Mch5 抗体
MCH5 抗体
xCaspase-8 抗体
zgc:92075 抗体
您还需要查找其他产品吗?
- Caspase 7 抗体
- Caspase 6 抗体
- Caspase 4 抗体
- Caspase 3 抗体
- Caspase 2 抗体
- Caspase 12 (Gene/pseudogene) 抗体
- Caspase 10 抗体
- Caspase 1 抗体
- CASP5 抗体
- CASP14 抗体
- CasLTR2 抗体
- CASKIN2 抗体
- CASK 抗体
- Casein Kinase 1, delta 抗体
- Casein Kinase 1 gamma 2 抗体
- Casein alpha-S2 抗体
- Casein alpha S1 抗体
- Casein 抗体
- CASC5 抗体
- CASC4 抗体
- Caspase 9 抗体
- Caspase Activity and Apoptosis Inhibitor 1 抗体
- CASQ2 抗体
- CASR 抗体
- CASS4 抗体
- CASZ1 抗体
- Catalase 抗体
- Catechol-O-Methyltransferase 抗体
- Cathelicidin 抗体
- Cathepsin 7 抗体
- Cathepsin B 抗体
- Cathepsin D 抗体
- Cathepsin E 抗体
- Cathepsin F 抗体
- Cathepsin G 抗体
- Cathepsin H 抗体
- Cathepsin K 抗体
- Cathepsin L1 抗体
- Cathepsin L2 抗体
- Cathepsin S 抗体



