CTNNB1 抗体
(Catenin (Cadherin-Associated Protein), beta 1, 88kDa (CTNNB1))
The protein encoded by this gene is part of a complex of proteins that constitute adherens junctions (AJs). AJs are necessary for the creation and maintenance of epithelial cell layers by regulating cell growth and adhesion between cells. The encoded protein also anchors the actin cytoskeleton and may be responsible for transmitting the contact inhibition signal that causes cells to stop dividing once the epithelial sheet is complete. Finally, this protein binds to the product of the APC gene, which is mutated in adenomatous polyposis of the colon. Mutations in this gene are a cause of colorectal cancer (CRC), pilomatrixoma (PTR), medulloblastoma (MDB), and ovarian cancer. Three transcript variants encoding the same protein have been found for this gene.[provided by RefSeq, Oct 2009].
Recommended next Filters:
247 results
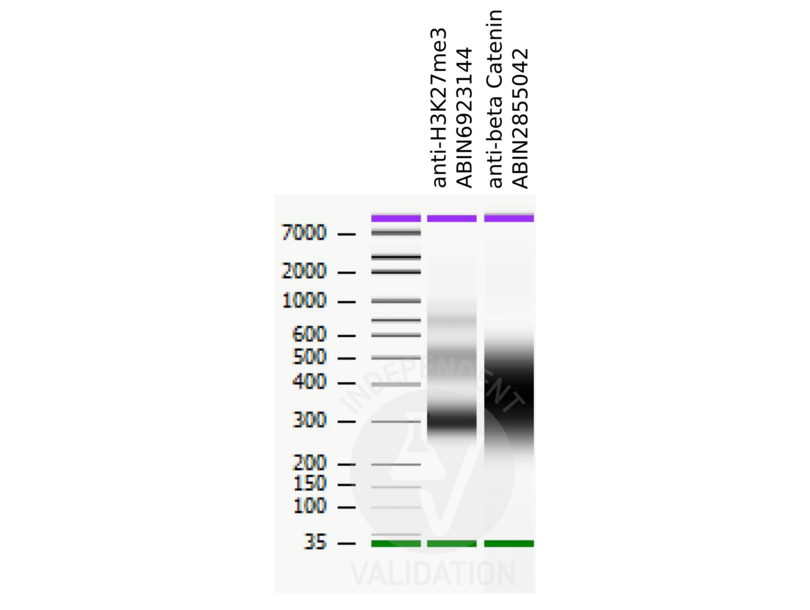


Independently Validated
Latest Publications for our CTNNB1 抗体
: "Wnt signaling activation induces CTCF binding and loop formation at cis-regulatory elements of target genes." in: Genome research, Vol. 35, Issue 8, pp. 1701-1716, (2025) (PubMed).
: "Exhaustive identification of genome-wide binding events of transcriptional regulators." in: Nucleic acids research, Vol. 52, Issue 7, pp. e40, (2024) (PubMed).
: "The β-catenin C terminus links Wnt and sphingosine-1-phosphate signaling pathways to promote vascular remodeling and atherosclerosis." in: Science advances, Vol. 10, Issue 11, pp. eadg9278, (2024) (PubMed).
: "Acute Intoxication With Alcohol Reduces Trauma-Induced Proinflammatory Response and Barrier Breakdown in the Lung via the Wnt/β-Catenin Signaling Pathway." in: Frontiers in immunology, Vol. 13, pp. 866925, (2022) (PubMed).
: "38" in: , Vol. 1363, Issue Nucleic acids research, pp. 28-39, (1991)
: "A new cut&run low volume-urea (LoV-U) protocol optimized for transcriptional co-factors uncovers Wnt/b-catenin tissue-specific genomic targets." in: Development (Cambridge, England), (2022) (PubMed).
: "Identification of candidate lncRNAs and circRNAs regulating WNT3/β-catenin signaling in essential hypertension." in: Aging, Vol. 12, Issue 9, pp. 8261-8288, (2020) (PubMed).
: "A novel c-Kit/phospho-prohibitin axis enhances ovarian cancer stemness and chemoresistance via Notch3-PBX1 and β-catenin-ABCG2 signaling." in: Journal of biomedical science, Vol. 27, Issue 1, pp. 42, (2020) (PubMed).
: "ARRDC3 suppresses colorectal cancer progression through destabilizing the oncoprotein YAP." in: FEBS letters, Vol. 592, Issue 4, pp. 599-609, (2019) (PubMed).
: "Progranulin protects the mouse retina under hypoxic conditions via inhibition of the Toll‑like receptor‑4‑NADPH oxidase 4 signaling pathway." in: Molecular medicine reports, Vol. 19, Issue 1, pp. 382-390, (2019) (PubMed).
: "Exhaustive identification of genome-wide binding events of transcriptional regulators." in: Nucleic acids research, Vol. 52, Issue 7, pp. e40, (2024) (PubMed).
: "The β-catenin C terminus links Wnt and sphingosine-1-phosphate signaling pathways to promote vascular remodeling and atherosclerosis." in: Science advances, Vol. 10, Issue 11, pp. eadg9278, (2024) (PubMed).
: "Acute Intoxication With Alcohol Reduces Trauma-Induced Proinflammatory Response and Barrier Breakdown in the Lung via the Wnt/β-Catenin Signaling Pathway." in: Frontiers in immunology, Vol. 13, pp. 866925, (2022) (PubMed).
: "38" in: , Vol. 1363, Issue Nucleic acids research, pp. 28-39, (1991)
: "A new cut&run low volume-urea (LoV-U) protocol optimized for transcriptional co-factors uncovers Wnt/b-catenin tissue-specific genomic targets." in: Development (Cambridge, England), (2022) (PubMed).
: "Identification of candidate lncRNAs and circRNAs regulating WNT3/β-catenin signaling in essential hypertension." in: Aging, Vol. 12, Issue 9, pp. 8261-8288, (2020) (PubMed).
: "A novel c-Kit/phospho-prohibitin axis enhances ovarian cancer stemness and chemoresistance via Notch3-PBX1 and β-catenin-ABCG2 signaling." in: Journal of biomedical science, Vol. 27, Issue 1, pp. 42, (2020) (PubMed).
: "ARRDC3 suppresses colorectal cancer progression through destabilizing the oncoprotein YAP." in: FEBS letters, Vol. 592, Issue 4, pp. 599-609, (2019) (PubMed).
: "Progranulin protects the mouse retina under hypoxic conditions via inhibition of the Toll‑like receptor‑4‑NADPH oxidase 4 signaling pathway." in: Molecular medicine reports, Vol. 19, Issue 1, pp. 382-390, (2019) (PubMed).
Aliases for CTNNB1 抗体
catenin beta 1 (CTNNB1) 抗体catenin (cadherin associated protein), beta 1 (Ctnnb1) 抗体
catenin beta 1 (Ctnnb1) 抗体
catenin (cadherin-associated protein), beta 1 (ctnnb1) 抗体
catenin beta 1 S homeolog (ctnnb1.S) 抗体
catenin beta 1 L homeolog (ctnnb1.L) 抗体
armadillo 抗体
beta-catenin 抗体
Bfc 抗体
Catnb 抗体
CHBCAT 抗体
CTNNB 抗体
ctnnb 抗体
CTNNB1 抗体
ctnnb1 抗体
id:ibd2058 抗体
Jup 抗体
LOC100217813 抗体
Mesc 抗体
MRD19 抗体
wu:fb73e10 抗体
wu:fi81c06 抗体
wu:fk25h01 抗体
您还需要查找其他产品吗?
You are here:



