Myc Proto-Oncogene protein 抗体
MYC 适用: 人 WB, ELISA, IF, IHC (p) 宿主: 小鼠 Monoclonal 1G7 unconjugated
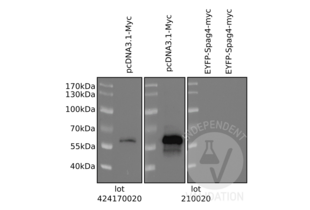
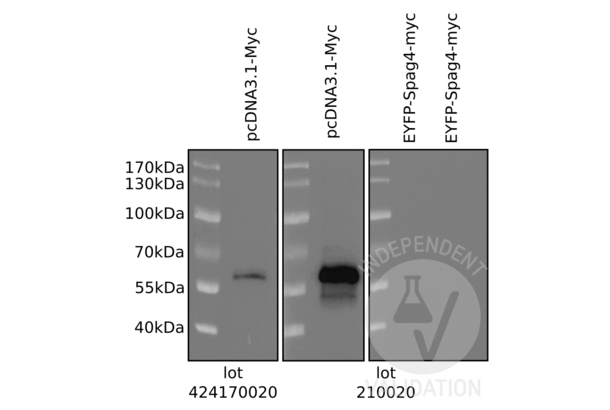 c-MYC 抗体 (AA 31-80) (ABIN1532205)
c-MYC 抗体 (AA 31-80) (ABIN1532205)
MYC 适用: 人, 小鼠, 大鼠 WB, IHC, ELISA, IF 宿主: 兔 Polyclonal unconjugated
Myc Proto-Oncogene protein 抗体 by Grade
Find Myc Proto-Oncogene protein 抗体 with a specific Grade. The Grade listed below are among those available. Click on a link to go to the corresponding products.
Myc Proto-Oncogene protein 抗体 by 适用
Find Myc Proto-Oncogene protein 抗体 for a variety of species such as anti-Human Myc Proto-Oncogene protein, anti-Mouse Myc Proto-Oncogene protein, anti-Rat Myc Proto-Oncogene protein. The species listed below are among those available. Click on a link to go to the corresponding products.
Myc Proto-Oncogene protein 抗体 by 抗体来源
Find Myc Proto-Oncogene protein 抗体 with a specific 抗体来源. The 抗体来源 listed below are among those available. Click on a link to go to the corresponding products.
Myc Proto-Oncogene protein 抗体 by 克隆形成能力
Find available monoclonal or polyclonal Myc Proto-Oncogene protein 抗体. Click on a link to go to the corresponding products.
Popular Myc Proto-Oncogene protein 抗体
- (1)
- (6)
- (5)
- (8)
- (3)
- (2)
- (7)
- (12)
- (12)
- (6)
- (2)
- (3)
- (4)
- (3)
- (3)
- (3)
- (3)
- (13)
- (2)
- (2)
- (2)
Latest Publications for our Myc Proto-Oncogene protein 抗体
: "Oxidative stress-induced mitophagy is suppressed by the miR-106b-93-25 cluster in a protective manner." in: Cell death & disease, Vol. 12, Issue 2, pp. 209, (2021) (PubMed).: "Long non-coding RNA CTSLP8 mediates ovarian cancer progression and chemotherapy resistance by modulating cellular glycolysis and regulating c-Myc expression through PKM2." in: Cell biology and toxicology, (2021) (PubMed).
: "Wnt/β‑catenin signaling modulates piperine‑mediated antitumor effects on human osteosarcoma cells." in: Molecular medicine reports, Vol. 21, Issue 5, pp. 2202-2208, (2021) (PubMed).
: "Manipulating ZmEXPA4 expression ameliorates the drought-induced prolonged anthesis and silking interval in maize." in: The Plant cell, Vol. 33, Issue 6, pp. 2058-2071, (2021) (PubMed).
: "ACSL4 promotes hepatocellular carcinoma progression via c-Myc stability mediated by ERK/FBW7/c-Myc axis." in: Oncogenesis, Vol. 9, Issue 4, pp. 42, (2020) (PubMed).
: "Cellular density-dependent increases in HIF-1α compete with c-Myc to down-regulate human EP4 receptor promoter activity through Sp-1-binding region." in: Pharmacology research & perspectives, Vol. 6, Issue 6, pp. e00441, (2019) (PubMed).
: "Neoisoliquiritigenin Inhibits Tumor Progression by Targeting GRP78-β- catenin Signaling in Breast Cancer." in: Current cancer drug targets, Vol. 18, Issue 4, pp. 390-399, (2018) (PubMed).
: "Sphingomyelin synthase 2 overexpression promotes cisplatin-induced apoptosis of HepG2 cells." in: Oncology letters, Vol. 15, Issue 1, pp. 483-488, (2018) (PubMed).
: "Upregulation of E‑cadherin expression mediated by a novel dsRNA suppresses the growth and metastasis of bladder cancer cells by inhibiting β-catenin/TCF target genes." in: International journal of oncology, Vol. 52, Issue 6, pp. 1815-1826, (2018) (PubMed).
: "Functional characterization of chloroplast-targeted RbgA GTPase in higher plants." in: Plant molecular biology, Vol. 95, Issue 4-5, pp. 463-479, (2018) (PubMed).
Aliases for Myc Proto-Oncogene protein 抗体
MYC proto-oncogene, bHLH transcription factor (MYC) 抗体v-myc myelocytomatosis viral oncogene homolog (LOC100136746) 抗体
nucleolar protein 3 (apoptosis repressor with CARD domain) (Nol3) 抗体
MYC proto-oncogene, bHLH transcription factor (Myc) 抗体
myelocytomatosis oncogene (Myc) 抗体
MYC proto-oncogene, bHLH transcription factor a (myca) 抗体
MYC proto-oncogene, bHLH transcription factor L homeolog (myc.L) 抗体
MYC proto-oncogene, bHLH transcription factor S homeolog (myc.S) 抗体
v-myc myelocytomatosis viral oncogene homolog (avian) (MYC) 抗体
MYC proto-oncogene, bHLH transcription factor b (mycb) 抗体
CG10798 gene product from transcript CG10798-RB (Myc) 抗体
anon-WO03040301.171 抗体
ARC 抗体
AU016757 抗体
B430311C09Rik 抗体
bHLHe39 抗体
bHLHe57 抗体
c-Myc 抗体
c-myc 抗体
C-MYC 抗体
c-MYC 抗体
c-myc II 抗体
CG10798 抗体
CMYC 抗体
cmyc 抗体
CMyc 抗体
D-Myc 抗体
d-myc 抗体
da 抗体
DM 抗体
Dm 抗体
dm 抗体
dm/dMyc 抗体
dm/myc 抗体
Dmel\CG10798 抗体
DMYc 抗体
Dmyc 抗体
dMYC 抗体
dMyc 抗体
dmyc 抗体
dMyc1 抗体
dmyc1 抗体
EG:BACN5I9.1 抗体
l(1)G0139 抗体
l(1)G0354 抗体
l(1)G0359 抗体
LOC100136746 抗体
mMyc 抗体
MRTL 抗体
MYC 抗体
myc 抗体
myc-B 抗体
myc1 抗体
Myc2 抗体
myc2 抗体
MYCC 抗体
Niard 抗体
Nird 抗体
NOP 抗体
Nop30 抗体
RNCMYC 抗体
zc-myc 抗体
zgc:55680 抗体



