TLR9 抗体
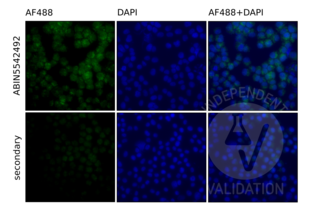
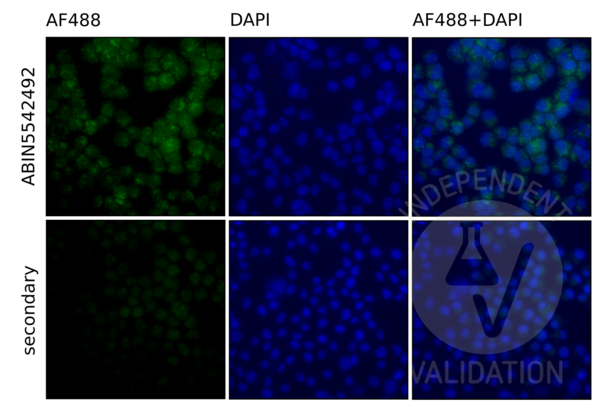 TLR9 抗体 (AA 868-1016) (ABIN5542492)
TLR9 抗体 (AA 868-1016) (ABIN5542492)
TLR9 适用: 人 WB, IHC, ELISA, FACS, ICC 宿主: 小鼠 Monoclonal 1B12H2 unconjugated
TLR9 适用: 人, 小鼠, 大鼠, 猴 WB, IHC, ELISA, FACS, IHC (p), ICC, IP, IF 宿主: 小鼠 Monoclonal 26C593-2 unconjugated
TLR9 抗体 by 抗体来源
Find TLR9 抗体 with a specific 抗体来源. The 抗体来源 listed below are among those available. Click on a link to go to the corresponding products.
TLR9 抗体 by 克隆形成能力
Find available monoclonal or polyclonal TLR9 抗体. Click on a link to go to the corresponding products.
TLR9 抗体 by 标记
Find TLR9 抗体 with a specific conjugate such as FITC, Biotin, PE. The conjugates listed below are among those available. Click on a link to go to the corresponding products.
Popular TLR9 抗体
- (7)
- (2)
- (8)
- (6)
- (3)
- (4)
- (4)
- (1)
- (2)
- (3)
- (7)
- (2)
- (3)
- (7)
- (2)
- (7)
- (2)
- (2)
- (2)
- (1)
- (1)
- (2)
Latest Publications for our TLR9 抗体
: "Differential expression patterns of Toll Like Receptors and Interleukin-37 between calcific aortic and mitral valve cusps in humans." in: Cytokine, Vol. 116, pp. 150-160, (2019) (PubMed).: "Co-expression of TLR-9 and MMP-13 is associated with the degree of tumour differentiation in prostate cancer." in: International journal of experimental pathology, Vol. 100, Issue 2, pp. 123-132, (2019) (PubMed).
: "A proliferation-inducing ligand (APRIL) induced hyper-production of IgA from tonsillar mononuclear cells in patients with IgA nephropathy." in: Cellular immunology, Vol. 341, pp. 103925, (2019) (PubMed).
: "Necroptosis in pulmonary macrophages mediates lipopolysaccharide-induced lung inflammatory injury by activating ZBP-1." in: International immunopharmacology, Vol. 77, pp. 105944, (2019) (PubMed).
: "Genistein modulates the expression of Toll-like receptors in experimental autoimmune encephalomyelitis." in: Inflammation research : official journal of the European Histamine Research Society ... [et al.], Vol. 67, Issue 7, pp. 597-608, (2018) (PubMed).
: "Elevated intragraft expression of innate immunity and cell death-related markers is a risk factor for adverse graft outcome." in: Transplant immunology, Vol. 48, pp. 39-46, (2018) (PubMed).
: "High Tidal Volume Induces Mitochondria Damage and Releases Mitochondrial DNA to Aggravate the Ventilator-Induced Lung Injury." in: Frontiers in immunology, Vol. 9, pp. 1477, (2018) (PubMed).
: "Toll-like receptors 2, 4 and 9 and hypoxia markers HIF-1alpha and CAIX in pancreatic intraepithelial neoplasia." in: APMIS : acta pathologica, microbiologica, et immunologica Scandinavica, Vol. 126, Issue 11, pp. 852-863, (2018) (PubMed).
: "Impaired Surface Expression of HLA-DR, TLR2, TLR4, and TLR9 in Ex Vivo-In Vitro Stimulated Monocytes from Severely Injured Trauma Patients." in: Mediators of inflammation, Vol. 2017, pp. 2608349, (2017) (PubMed).
: "High toll-like receptor (TLR) 9 expression is associated with better prognosis in surgically treated pancreatic cancer patients." in: Virchows Archiv : an international journal of pathology, Vol. 470, Issue 4, pp. 401-410, (2017) (PubMed).
Aliases for TLR9 抗体
toll like receptor 9 (TLR9) 抗体toll-like receptor 9 (Tlr9) 抗体
CD289 抗体



